|

"Cinco
sentidos, una perspectiva
de la reproducción de las orquídeas"
por
Luis Mendoza Teytud
Colaboró
- Katia Seagull
La
nariz humana contiene en su interior tejido que posee receptores que
responden a estímulos provocados por las moléculas de substancias volátiles
y que son capaces de transformar esa señal química en
impulsos eléctricos que viajarán hasta llegar a distintas zonas
del encéfalo, posterior a lo cual se genera una respuesta por parte del
individuo. En su
ruta, esos impulsos atraviesan por estructuras neurológicas
relacionadas con la memoria, las emociones, los sentimientos, etc., es
por este motivo que el sentido del olfato está relacionado íntimamente
con esos factores. Nuestro olfato es, por sus dimensiones y
por el número de receptores y las características propias de éstos últimos,
probablemente uno de los menos favorecidos por la naturaleza, superado
por muchos otros mamíferos y tal vez por otros animales, incluidos los
insectos. En estos últimos, los sensorios olfativos se localizan
principalmente en las antenas pero pueden estar dispuestos en otras
zonas corporales y ser capaces de discernir entre aromas comunes y
aquellos relacionados específicamente con las feromonas, sustancias
secretadas o excretadas capaces de despertar una respuesta social o sistémica
determinada en individuos de la misma especie. En el tema que nos ocupa
el papel de las feromonas sexuales es crucial.
Para
nuestro olfato, mas o menos el 75% de las orquídeas poseen
fragancia, la mayor parte de ellas son epífitas pero existen también
algunas especies terrestres que tienen esta característica. Es muy
probable que los polinizadores perciban la presencia de moléculas de
sustancias volátiles en cantidades infinitamente pequeñas y distantes,
según algunos autores, hasta de una decena de kilómetros.
Esta es una tarea materialmente imposible para un sentido del
olfato tan limitado como el del ser humano.
En algunas referencias se hace énfasis también a las grandes
ventajas que para los insectos representa el tener en sus antenas además
los receptores del tacto y del gusto, es decir reúnen en áreas
diminutas varios de sus sentidos.
Interrelación
flor – polinizador
En el siglo antepasado, Delpino propuso el término de síndrome de
polinización o lo que actualmente algunos autores llaman simplemente síndrome
floral, para referirse a aquel conjunto de características como color,
tamaño, forma, producción de perfumes o néctar, época de floración,
etc., gracias a las cuales atraen a determinado polinizador.
Si bien es un término que se acuño hace más de
cien años y que en la actualidad
no es aceptado totalmente, desde el punto de vista didáctico
resulta útil. Esto no
significa que afirmemos que las
características de determinada especie impliquen que sea exclusivamente
el polinizador el que “tenga derecho” de posarse en ella.
Las flores de una sola especie pueden ser visitadas por colibríes,
escarabajos, abejas euglosinas, etc. pero eso no implica el que todos
puedan polinizarla o sólo algunos de ellos o solamente uno de ellos.
En otras palabras, en muchas plantas ( no orquídeas) las flores
pueden tener varios vectores, por tanto, generalmente sus características
no son específicas para determinado polinizador ficas
para determinado polinizador acterolinizadores, por tanto el sis puedan
polinizarla, solo algunos de ellos o solamente uno. En el caso
particular de las orquídeas para que ocurra la polinización sí se
requiere de un vector específico que sea capaz de desplazarse, que
pueda ascender en el aire, que se comporte de determinada manera y que
tenga las características físicas necesarias para remover y depositar
el polinario de y en determinado sitio. En la familia Orchidaceae el
vector debe ser entonces un elemento vivo, por lo que la polinización
es biótica, ese polinizador corresponde siempre al reino animal, por
tanto, la polinización es zoofílica y muchas de ellas específicamente
entomófila por ser polinizadas por insectos y más específicamente
sarcófila o coprosarcófila ( polinización por moscas), psycófila (
por mariposas), melitófila (por abejas), etc. u ornitófila ( por
aves).
|
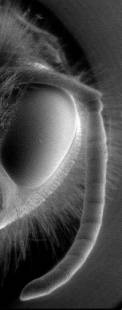
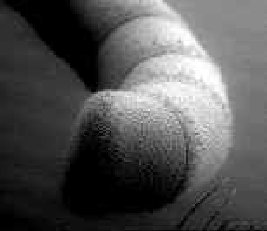
Cuadro
1 -
Microfotografía panorámica
de la antena de alguna abeja. °La
imagen de
la derecha
corresponde a un acercamiento de la antena, la del centro
es un acercamiento de la punta de la antena, cada pequeña
vellosidad contiene terminaciones nerviosas responsables del
olfato, entre otros sentidos.
|
|
|

|
|
Los cambios
evolutivos, entre las flores y polinizadores, son considerados por
algunos estudiosos como una de las facetas del fenómeno conocido como
coevolución. Las modificaciones en las características morfológicas y
fisiológicas de la flor y/o el polinizador han ido adaptándose de tal
modo que “uno es complemento del otro”. En otras palabras, para
muchos autores, en las orquídeas, la definición
del proceso podría ser completa: “la evolución conjunta de
dos o más especies que no se cruzan entre sí pero que poseen una
estrecha relación ecológica, a través de presiones de selección recíprocas,
la evolución de una de las especies que forman esta relación es
parcialmente dependiente de la evolución de la otra” El vínculo que se establece en este caso entre orquídea y
polinizador se conoce como mutualismo y consiste en que, tanto una parte
como la otra, resultan
beneficiados aunque en algunos reportes prefieren nombrar a esta relación
como explotación mutua. No se debe pasar por alto que en el caso
particular de las orquídeas el mutualismo resulta relativo porque la
coevolución ha llevado a resultados tan sofisticados como el engaño,
de tal modo que en algunos casos la única que resulta beneficiada
en la relación es la flor puesto que como consecuencia del engaño
resulta polinizada. Para
algunos autores la coevolución orquídea – polinizador no existe y el
mutualismo surgió más bien como una adaptación unilateral por parte
de las orquídeas, ante los polinizadores que habrían aparecido con
anterioridad y que tendrían ya adaptaciones con otras plantas
productoras de flores.
El mutualismo
orquídea – polinizador ha llegado a tal extremo que los polinizadores
suelen ser bastante específicos para las especies. Si bien esto
garantiza el que la inmensa mayoría de las veces el polen de una flor
vaya a dar al estigma de otra de la misma especie y no haya
“desperdicio de material genético”, representa también una
limitante porque en ausencia del polinizador no habrá reproducción
sexual y en ausencia de las flores probablemente los insectos tendrían
cierta limitación en la posibilidad de cortejar y aparearse, como
veremos más adelante. Un ejemplo concreto en la especificidad del
polinizador es Vanilla planifolia que es polinizada en la naturaleza
exclusivamente por una abeja del género Melipona, cuyo tamaño y
características físicas son perfectas para que el pequeño insecto,
atraído por las características de las flores sea el polinizador único
de esa especie. Es por este
motivo que la polinización de la vainilla no ocurre fuera de su hábitat
natural en México puesto que es la única parte en donde aquel insecto
existe.
Algunas de las
especies con polinización más sofisticada son las Angraecoides, como
Jumella, Angraecum, Aeranthes, etc. Estas plantas producen flores
blancas con fragancias fuertes que son mas intensas por la noche.
Atraen así a insectos de hábito nocturno, polillas de distintas
especies. Angraecum
sesquipedale, posee un nectario tan profundo que solamente es alcanzado
por la polilla de la especie Xanthopan morgani praedicta cuyo probóscide
rebasa los 25 cms y que fue descubierta hasta finales del siglo XIX,
posterior a la muerte de Darwin y de otros investigadores, cuya atención
se centró por mucho tiempo en la sesquipedale y que argumentaban ya la
posibilidad de que ese insecto existiese, motivo por el que fueron
criticados por los entomólogos de la época.
Existen otros géneros
de flores blancas que recompensan también con néctar y son
polinizadas por la noche, es decir comparten un síndrome floral
muy parecido con las angraecoides, dos ejemplos son especies de
Brassavola y Aerangis , muchas de ellas huelen a madreselva, jazmín o
gardenia.
Los síndromes
florales no son necesariamente exclusivos de las orquídeas, es decir,
algunas flores de estas tienen color, olor, forma, etc. parecidas a las
floraciones de otra familia de plantas no relacionadas y pueden ser
polinizadas por el mismo insecto, comparten el mismo síndrome floral.
Los polinizadores, al forrajear* asocian esas características de las
flores con determinada recompensa de alimento; la orquídea es entonces
polinizada por engaño. En esta variante resultan atraídos
polinizadores machos y hembras puesto que no hay ningún elemento sexual
en la ecuación . Esta es considerada, en plantas la forma mulleriana de
mimetismo. Podría decirse que en este caso para la orquídea “es
bueno tener flores con características similares a la de otras plantas
del entorno”. Es probable que más o menos 10 mil especies de la
familia Orchidaceae sean polinizadas con esta estrategia, en la que
participan abejas, polillas, mariposas, moscas y escarabajos.
* Este término no figura en el diccionario de la real academia de la
lengua, pero se usa para referirse a la búsqueda colección de
alimentos por aves e insectos.
En algunas orquídeas
las flores no solamente son morfológicamente parecidas a la de las
hembras de algunos insectos sino que, además,
su fragancia es muy parecida a las de sus feromonas sexuales. El
género Ophrys, específicamente la especie insectífera, por ejemplo,
produce flores con características parecidas a las de las hembras de
las avispas Argogorytes mistaceous . Por si fuera poco, se sabe que esta
flor, apenas abre, produce
la feromona que atrae a los machos de esa especie y, si posterior a
varios días no es polinizada, produce feromonas que atraen a los machos
de Argogorytes campestris. Forma,
color y aroma son, entonces, un atractivo irresistible desde el punto de
vista reproductivo y los
machos, en el intento de
aparearse con la supuesta hembra, desprenden el polinario que será
llevado a otra flor . Este es un engaño total,
la flor a los ojos de los machos es una hembra, ni siquiera
parecen relacionarla con una flor, es un engaño sexual que
explota el instinto reproductivo de los machos.
Para algunos autores, el engaño mediante la copia de un modelo
“no planta”, por parte de la orquídea se llama mimetismo batesiano
:” Es valioso ser distinto a las flores del entorno”.
El caso de las Dracula es particular: Se sabe que son polinizadas por
pequeñas moscas drosofiloides
del género Zygothrica. Estos insectos cortejan, se aparean
y probablemente depositen sus huevecillos en hongos que comparten
hábitat con ese genero de orquídeas.
El labelo de las Dracula es parecido en
color, forma y olor a esos hongos. Aquí
el engaño está relacionado con la conducta sexual, la orquídea atrae
a ambos géneros que durante su rito sexual polinizan a las Dracula.
Es esta otra expresión del mimetismo batesiano, el hongo que “
sirvió como modelo” para el labelo no es una flor, ni siquiera es una
planta. Hay artículos en los que citan que este modo de polinización
podría ser de tipo atracción “rendezvous”, tipo refugio o incluso
mimetismo mulleriano ( ver más adelante ). Habría que valorar
cuidadosamente, tal parece que para otros investigadores, en la atracción
“rendezvous”, la orquídea imita a otra flor del entorno, es decir a
una planta, no a un hongo y en la polinización relacionada con la búsqueda
de refugio la mayor parte de las veces se describe solamente como aquel
modo de polinizaición que ocurre cuando el vector busca abrigo, sin
distinguir hembras ni machos y sin que implique
cortejo ni apareamiento. Para
algunos autores el mimetismo batesiano podría ser responsable de la
polinización de mas o menos una tercera parte de todas las especies de
orquídeas.
Otro
modo de engaño relacionado con la conducta sexual de los machos
y la búsqueda de alimento por parte de las hembras es el que se conoce
como atracción “rendezvous”: Algunas especies de orquídeas
europeas y africanas florecen simultáneamente con otras plantas no
relacionadas y con las que comparten características. Estas últimas
proporcionan polen y néctar a las hembras de determinadas especies de
abejas. Los machos frecuentan a estas flores puesto que es tiempo en que
sus hembras están en pleno forrajeo. Hembras y machos, resultan engañados,
se han posado en una orquídea. En el proceso cortejo-apareamiento
ocurre la polinización. Para
algunos autores la atracción “rendezvous” es una forma de mimetismo
mulleriano puesto que la flor “modelo” tiene gran parecido con la
orquídea, sin embargo en este caso son las hembras las que resultan
primeramente atraídas ante la posibilidad de conseguir forraje,
mientras que los machos están realmente en búsqueda de hembras, no de
alimento.

La similitud de
algunas especies de Tolumnia y Oncidium
con himenópteros favorece que el movimiento de las flores por acción de las
corrientes de aire, hace que los insectos al defender su territorio
“ataquen a la flor” y la polinicen.
La orquídea resulta polinizada y el insecto con un entrenamiento
mayor para defender su zona. Parece ser una situación que se ve raras
veces y que no es selectivo de hembras ni machos.
En algunas orquídeas del mediterráneo como las Serapia, las flores de
color rojo intenso tienen un largo tubo cuya entrada semeja la de un
nido de algunas abejas y su interior ofrece resguardo con temperatura
hasta 3°C por arriba de la ambiental. Resulta entonces un excelente
resguardo para los polinizadores que, en el proceso entrada – salida,
polinizan. En este caso la
estrategia del engaño suele no ser aceptada por muchos autores, la
relacionan simplemente con el resguardo que instintivamente busca el
insecto.
En los casos de
mimetismo relacionado con el engaño en busca de alimento y con el
pseudoapareamiento, se ha demostrado que los polinizadores con relativa
facilidad pueden aprender a diferenciar olores y/o colores entre la flor
modelo ( a la que la orquídea imita),
de la “copia” (la orquídea que imita).
Se podría decir que son formas agotables de mimetismo, que se
repiten cuando los polinizadores “que aprendieron” se trasladan a
otras zonas con colonias de la misma especie de orquídeas o con la
descendencia inexperta de esos polinizadores.
Se ha visto que el depósito de polen de una flor a otra, de la
misma planta (polinización a la que se conoce como geitenogamia, que
genéticamente corresponde a un tipo de autopolinización) se ve
raramente porque el polinizador evita, toda vez que ha sido engañado,
posarse en otra flor de la misma inflorescencia o planta.
Recorre, entonces, una distancia más o menos larga hasta que se
encuentra otra colonia de orquídeas
en donde nuevamente será burlado y depositará el polen que lleva de su
trampa anterior. Es
frecuente que cuando se produce geitenogamia los frutos se aborten en
etapas tempranas o los embriones se desarrollen solo precariamente.
El
particular caso de las
abejas
Euglossini
Las abejas euglosinas pertenecen a la tríbu Euglossini, que comprende
cinco géneros y alrededor
de 200 especies que forman
parte del grupo de las abejas corbiculadas. Estos insectos tienen una
relación muy estrecha con las orquídeas, (especíalmente con epífitas)
a tal grado que se les ha llamado “orchid
bees" o “las abejas de las orquídeas”. Se pueden ver desde México
hasta Argentina y el Caribe. Se conocen dos ancestros fósiles de ellas,
fueron encontrados en República
Dominicana y se sabe que existieron hace quince a cuarenta y cinco
millones de años.
Los machos de
abejas euglosinas, atraídos por el perfume, se posan sobre las flores
de orquídeas (generalmente de las subtribus Stanhopeinae, Catasetinae,
Diacheinae, Zygopetalinae, algunos géneros de Oncidiinae y
Cyrtopodiinae). Con sus antenas exploran y al encontrar el sitio en
donde se producen las substancias responsables de la fragancia
(sequiterpenos y sus derivados, por ejemplo), secretan en sus labios
sustancias grasas con las que hacen una mezcla que recogen
con sus patas delanteras, se elevan y despliegan sus patas
medias, sobre las que frotan las delanteras, con el consecuente
desplazamiento de aquella mezcla. Finalmente, frotan las patas medias contra la porción tibial
ensanchada de las patas posteriores, en donde se encuentra tejido
espongiforme en donde se acumula el material. Es probable que en ese
sitio de las patas haya glándulas productoras de determinadas
sustancias que completan la fragancia o feromona necesaria para el
cortejo. Durante el proceso del arrastre de la fragancia,
el polen es desprendido por la abeja y llevado en su dorso a otra
flor.Como se describe adelante, en casos concretos los sensores de la
flor disparan el polinario. Para cortejar, el insecto macho acomoda
sus extremidades con una disposición espacial que permite que
aquella estructura quede expuesta y libere el perfume que teóricamente
atraerá a las hembras . En algunas especies, los machos de Euglossini
tienen una anatomía más sofisticada porque poseen una superficie
aterciopelada en la porción tibial de sus patas medias.
Colectan ahí temporalmente sustancias volátiles que esparcen
durante el vuelo mediante la vibración, a modo de ventilador, de alas
auxiliares. Estas
estructuras son rudimentarias en las hembras.
Si
el perfume de las flores fuese la feromona en si, seguramente las
hembras también resultarían
atraídas, no solamente los machos. Darwin y otros autores del siglo XIX hacían ya alusión
a la importancia de las fragancias de las orquídeas pero no tenían un
concepto claro de su papel en la naturaleza.
Algunos científicos relacionaron la presencia de machos en la
flores con la posibilidad de que su olor fuese el propio de las
feromonas producidas por las hembras puesto que observaron como los
machos “rascaban” con las vellosidades de sus patas delanteras los pétalos
y sépalos y minutos después quedaban quietos, “como embriagados”
por la frustración. Otra
teoría aún más antigua y errónea citaba la posibilidad de que las
abejas euglosinas acudieran a las flores para alimentarse.
Aquellas
flores que producen fragancias que atraen a los machos Euglossini forman
un grupo al que, por su importancia y por ser de los más estudiados se
ha llamado “Síndrome de polinización euglosino” y está formado
por alrededor de 650 especies de orquídeas que son polinizadas
exclusivamente por esos insectos.
Las
doscientas especies de Euglossini se alimentan del polen y néctar de otras flores, de hecho la
fuente de energía de muchos otros polinizadores de orquídeas se basa
en los productos de otras plantas. Algunas orquídeas ofrecen algo de néctar
como recompensa, pero su polen nunca es un alimento. Hay géneros como Poslistachya poseen pseudopolen y falsas
anteras, otras tienen vellosidades y papilas, ambas ricas en almidón y proteínas que son colectados por los
polinizadores.
Se
ha demostrado que muchas especies euglosinas pueden buscar
determinada fragancia, es decir la misma especie de orquídea
pude ser visitada por varias especies de euglosina.
Sin embargo, el que una de ellas se pose en determinada orquídea
no significa necesariamente que será el vector, se requiere que sea
exclusivamente aquella especie que tenga el tamaño y el comportamiento
idóneo para desprender y posteriormente depositar el polen.
|
|
|
|
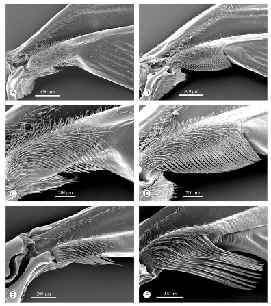
Cuadro
2 - °Las imágenes
del lado izquierdo muestran los apéndices de las alas
secundarias de algunas especies de Euglosinii, las de la derecha
corresponden los machos de las mismas. Se puede observar la
diferencia en dimensiones. |
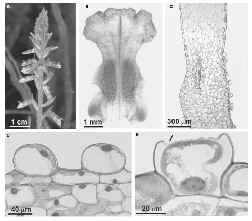
Cuadro
3 -
°Ciclopogon elatus. La imagen A muestra la vara floral. En B se ha
disecado el labelo, las dos áreas simétricas más obscuras
corresponden a los osmóforos en forma de vellosidades. C
corresponde a un corte longitudinal de la vellosidad.
D muestra las células de una de las vellosidades de un
capullo. En E se
observa una célula de la vellosidad con una depresión en su
superficie, lo que significa que está en plena liberación de
sustancias volátiles. |
En
los géneros Mormodes, Cycnoches y Catasetum de la subtribu Catasetinae
existe una adaptación peculiar: Sus flores masculinas poseen
estructuras extremadamente sensibles que al menor estímulo por parte
del macho de euglosina expelen el polinario con velocidades hasta de
alrededor de 3 metros por segundo, es decir casi 11 Km/hora. De ese modo
el polinario, por medio de su corpúsculo, y gracias a sus cojines
pegajosos queda firmemente adherido al dorso del insecto.
Nótese que en
el caso de la polinización por machos Euglossini
no hay engaño. Si bien hay relación con la conducta sexual, el
insecto colecta sustancias volátiles pero ni hay pseudoapareamiento con
la flor, ni las hembras acuden a ese sitio.
Es interesante también tener en mente que las euglosinas
colectan sustancias volátiles de otras familias de plantas
(cuyas
flores también polinizan). Es decir, no dependen exclusivamente de las
orquídeas. Además, no solo colectan sustancias de las flores, lo hacen
también de madera en descomposición, hongos, etc.
Una observación
interesante en prácticamente todas las orquídeas, independientemente
del modo en que se logre el cruce, es que los polinizadores habitualmente procuran las flores más
grandes, las de mejor forma y las de mejor colorido.
Aspectos
importantes de las fragancias
Los
perfumes de las orquídeas se producen en células esparcidas en toda la
epidermis de las flores o en estructuras glandulares llamadas osmóforos,
órganos localizados en pétalos y sépalos y, en algunas especies,
confinadas exclusivamente al labelo, seguramente para atraer directamente a los polinizadores al espacio crítico que queda entre ese
pétalo modificado y la columna.
La actividad productora de fragancia
por parte de los osmóforos cambia según  la hora del día, de acuerdo a la especie, aún
cuando las flores están abiertas las 24 horas. Los niveles altos de
humedad y temperatura elevada pueden incrementar el metabolismo de las
glándulas, lo cual se traduce en una acentuación de las fragancias. La cantidad de susbstancias volátiles que
producen las orquídeas son pequeñas puesto que de otro modo
resultarían tóxicas para la planta, se sabe que las que producen mayor
volúmen son miembros de los géneros Stanhopea, Herschelia y Catasetum.
Además, el proceso representa gasto extra de energía, probablemente
por eso el comportamiento sea cíclico y su presencia coincida con el
momento en que merodean los polinizadores.
Sin embargo, hay
orquídeas que tienen dos ciclos,Clowesia rosea huele a mentol por la mañana
y a canela por las tardes; Catasetum expansum por la mañana a
trementina y por la noche a pan de centeno.
En otros casos la producción es las 24hs, con
momentos pico por las noche, como sucede en Epidendrum difforme.
Nuestro olfato, y probablemente el de los polinizadores, pueden captar
los aromas cuando la flor tiene determinado grado de madurez,
seguramente no en flores jóvenes ni en regresión.
Dependiendo de la especie o de las especies que dieron lugar a
determinado híbrido, la intensidad del perfume puede acentuarse
temprano por la mañana, al medio día o por la noche.
Las flores de las plantas sanas son especialmente perfumadas. la hora del día, de acuerdo a la especie, aún
cuando las flores están abiertas las 24 horas. Los niveles altos de
humedad y temperatura elevada pueden incrementar el metabolismo de las
glándulas, lo cual se traduce en una acentuación de las fragancias. La cantidad de susbstancias volátiles que
producen las orquídeas son pequeñas puesto que de otro modo
resultarían tóxicas para la planta, se sabe que las que producen mayor
volúmen son miembros de los géneros Stanhopea, Herschelia y Catasetum.
Además, el proceso representa gasto extra de energía, probablemente
por eso el comportamiento sea cíclico y su presencia coincida con el
momento en que merodean los polinizadores.
Sin embargo, hay
orquídeas que tienen dos ciclos,Clowesia rosea huele a mentol por la mañana
y a canela por las tardes; Catasetum expansum por la mañana a
trementina y por la noche a pan de centeno.
En otros casos la producción es las 24hs, con
momentos pico por las noche, como sucede en Epidendrum difforme.
Nuestro olfato, y probablemente el de los polinizadores, pueden captar
los aromas cuando la flor tiene determinado grado de madurez,
seguramente no en flores jóvenes ni en regresión.
Dependiendo de la especie o de las especies que dieron lugar a
determinado híbrido, la intensidad del perfume puede acentuarse
temprano por la mañana, al medio día o por la noche.
Las flores de las plantas sanas son especialmente perfumadas.
Las
orquídeas producen 50 o más diferentes esencias florales. Muchos de
sus componentes han sido identificados en los últimos 20 – 30 años.
El número
de las substancias que pueden ser producidas y su efectividad varía según
las especies.
Muchas de
las esencias producidas por las orquídeas son simples componentes aromáticos,
tales como alfa y beta -pinene, myrcene, alfa -phelandrene, cineole,
ocimene, p-cymene, citronellae, linalool, geraniol, methyl benzoato,
beta -terpineol, benzyl acetato, perperitone, d-carvone, methyl
salicylato, nerol, 2- phenylethyl acetato, 2- phenylethanol, methyl
cinamato, eugenol y skatol.
La
efectividad de los componentes estructuralmente similares puede variar.
Por ejemplo, 300 abejas euglosinas
macho fueron atraídas por muestras de eugenol, pero sólo una
fue hacia el methyl isoeugenol.
Algunas substancias naturales afines a aquellas producidas por
orquídeas no son muy atractivas para las abejas. En cambio, ciertos
componentes que no ocurren en la naturaleza
o no están presentes en las esencias de orquídeas (anisyl
acetato
por
ejemplo, o el alfa- y beta -ionone, no el methyl ionone) son buenos
atractivos para las abejas.
Muchas de
las esencias producidas por las orquídeas son fragancias que atraen a
las abejas y otros vectores polinizantes. Ellas son también agradables
para los humanos, lo cual es el motivo de la popularidad de ciertas
especies, por ejemplo algunas cattleyas, Stanhopeas, etc.
Ciertos
olores, por ejemplo el skatol, son decididamente desagradables, como lo
indica su nombre, y atraen a vectores que usualmente son atraídos por
materia en descomposición. Tal el caso de Batemannia lindenii, de
Venezuela, el Bulbophyllum
de Bogor, o la Masdevalia foetens; de México Brassia verrucosa y
Encyclia radiata.
Composición
de las fragancias
de algunas orquídeas
Brassavola
digbyana Lindl. -
citronellol
Brassavola
perrinii Lindl.– alfa -
pinene, beta - pinene, 1,8- cineole
Catasetum
macrocarpum L.C.Rich.–
alfa - pinene, beta - pinene, 1,8 cineole.
Catasetum
pileatum Rchb.f. - alfa -
pinene, beta -pinene, 1,8 cineole, methyl benzoato, benzyl acetato,
carvone.
Stanhopea
tigrina Baterm. Ex Lindl.-
benzyl acetato.
Para
los humanos, el encuentro con las fragancias de las flores y en este
caso de las orquídeas, evidentemente nada tiene que ver con aspectos
reproductivos o de alimentación pero cuando esta característica es
agradable suele ser relevante. Por
su aroma, estas flores han sido llevadas
hasta escenarios internacionales con la asistencia de expertos en
perfumes y en el cultivo de orquídeas, por ejemplo el concurso de
fragancias implementado en la Muestra Internacional de Japón en 1989 y
en los Estados Unidos por parte de la Greater New York Orchid Society en
1992 .
°
Las imágenes con este símbolo fueron tomadas de algunos sitios libres
de la red, su autoría no es nuestra.
Luis Mendoza Teytud
-
México -

|